
Introduction
Facteurs héréditaires permettant de transmettre des caractères à la descendance.
I. Gènes et caractères
Caractère : ce qu’on observe sur un individu (couleur…). Les caractères sont contrôlés par des facteurs cellulaires qui génèrent ces caractères : les gènes.
I.A. Mise en évidence de l'existence de l'information génétique (expérimental)
I.A.1. Observation de cultures de bactéries
I.A.1.a. Pourquoi des observations sur les micro-organismes ?
Parce qu’ils ont des caractéristiques qui en font un matériel de choix :
- Ils se reproduisent vite, les générations se succèdent rapidement (1/30 minutes) ;
- La descendance est nombreuse et permet les traitements statistiques ;
- Les cultures prennent très peu de place (peu coûteux) ;
- Ce sont des organismes haploïdes : les gènes ne sont présents qu’en un seul exemplaire. Le gène s’exprime donc tel qu’il est. Chez l’homme, la diploïdie génère des phénomènes de dominance, de récession…
I.A.1.b. Un exemple de résultats de culture
Echerichia Coli (colibacile). Il existe des souches sauvages et des souches mutantes. La souche sauvage est prototrophe (une seule source de carbone, le glucose). A partir de carbone, la cellule fabrique toutes les autres molécules organiques. Le glucose est aussi la source d’énergie.
Il existe des souches auxotrophes qui ne peuvent pas utiliser que le glucose. Elles ont besoin de compléments. Exemple souche Tryptophane-, Adénine-, Histidine--. Elle se développe sur un milieu de culture complémenté en Tryptophane, Adénine, Histidine.
- Souche autotrophe : Besoins = eau, sels minéraux et glucose = Milieu minimum M0
- Souche auxotrophe : Besoins = eau, sels minéraux, glucose, Tryptophane, Adénine, Histidine = Milieu complémenté = M0 + Try + Ad + Hist
Etude de transmission de caractères métaboliques : La souche auxotrophe possède un défaut métabolique qui l’empêche de fabriquer Try, Ad et Hist. On va étudier le caractère « aptitude à fabriquer du Tryptophane ». Il y a 2 phénotypes différents :
- Phénotype Tryptophane+ capable de le fabriquer
- Phénotype Tryptophane- incapable de le fabriquer
I.A.1.c. Interprétation des résultats
La colonie se développe, par division des cellules. Apparaissent de nouvelles générations de bactéries qui possèdent les mêmes caractéristiques métaboliques que les générations mères. Les caractères métaboliques se transmettent d’une génération à l’autre. Ces caractéristiques sont transmissibles. Donc il existe dans la cellule des facteurs héréditaires capables de se transmettre de génération en génération et qui génèrent les caractères : ce sont les gènes.
1 gène → 1 caractère
On constate cependant 2 anomalies :
Boite 7 : apparition d’une colonie. La nouvelle génération est capable de fabriquer de l’adénine. Son information génétique s’est modifiée au cours des générations. Elle a récupéré le gène Ad+. Le gène de l’adénine a été modifié : il y a eu mutation. Ces gènes peuvent donc être modifiés.
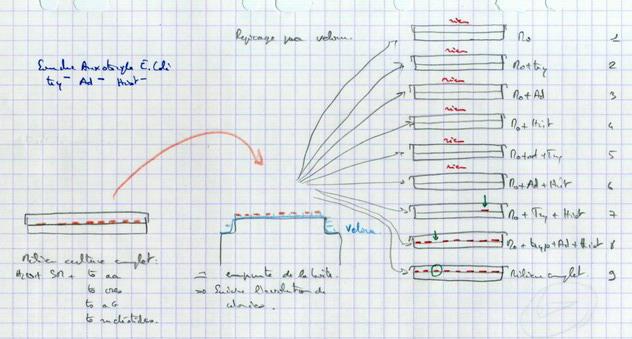

Boite 8 : 1 colonie est absente. Elle a eu une exigence supplémentaire que n’avaient pas les générations précédentes. Il y a eu modification de l’information génétique. 1 gène a été muté faisant apparaître une modification des exigences métaboliques. On peut rechercher l’élément qui lui manque (cuisine). Le protocole expérimental est le suivant : on opère un crible de sélection (repiquage sur velours) qui permet de sélectionner de nouvelles souches. On obtient des clones : toute la population descend d’une unique bactérie. Ici, on obtient 2 nouveaux clones.
I.B. Comment les gènes contrôlent-ils les caractères ?
Le gène est un facteur cellulaire. Le caractère est un facteur observable. Comment un gène peut-il contrôler l’apparition d’un caractère ? Le premier concept date de 1948.
1 gène → 1 π (Beadle et Tatum, 1948)
I.B.1. Le contexte scientifique
| Ils travaillaient sur un champignon, Neurospora, avec des souches sauvages et des souches auxotrophes (arginine-) cultivées sur un milieu complémenté en arginine. Ces travaux ont conduit à la découverte de la voie de biosynthèse de l’arginine. | |
Pour les souches Arginine-, une des étapes ci-dessus n’est plus possible. Impossibilité d’effectuer 1 ou les 3 étapes ? Si une seule est bloquée, de laquelle s’agit-il ? Tous les mutants auxotrophes sont-ils identiques ? |
|
I.B.2. Tous les mutants Arg- sont-ils identiques ? Non
I.B.2.a. Mise en évidence de l'existence de 3 groupes de mutants auxotrophes
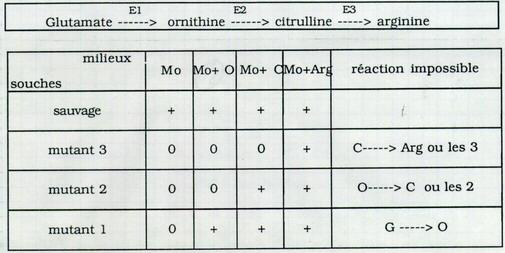
Tous les mutants auxotrophes Arg- ne sont pas identiques bien qu’ils soient identiques phénotypiquement. Il y a 3 groupes de mutants auxotrophes différents qui ont des potentialités différentes.
Hypothèse : dans chacun des cas, une seule étape n’était pas réalisée car il manquait l’enzyme correspondante.
| 1 gène conditionne l’apparition d’une enzyme | ||
| 1 gène | → 1 enzyme | |
| dans ce cas | 3 gènes | → 1 caractère |
Plusieurs gènes sont nécessaires à l’apparition d’un caractère visible.
I.B.2.b. Confirmation de l'absence d'une enzyme dans chacun des groupes de mutants par le test d'activité enzymatique
Broyage des mutants : ils gardent le surnageant. Dans des tubes à essai, ils recherchent les enzymes en ajoutant le substrat et en regardant s’il disparaît et s’il est transformé en produit.
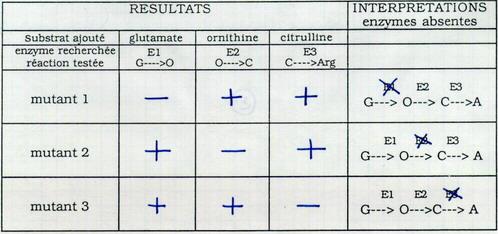
Donc 1 gène contrôle 1 enzyme. Ceci est vérifié par des tests biochimiques. Un gène est une sous unité du matériel héréditaire. Ce gène contrôle l’apparition d’une protéine dans la cellule. Le caractère n’est généralement réalisé que grâce au concours de plusieurs gènes. Dans une cellule, un gène existe sous deux allèles différents au minimum. On ne sait toujours pas ce qu’est un gène (nature, support…).
I.C. Quand on ne connaît pas les voies métaboliques, comment détermine-t-on si un caractère est sous contrôle de plusieurs gènes ?
Grâce au test de l’allélisme complémentation. Pour un mutant auxotrophe pour X :
- Tous les mutants sont-ils identiques ?
- Y a-t-il des étapes métaboliques intermédiaires ?
I.C.1. Principe général du test d'allélisme complémentation
Ils ne peuvent être réalisés que sur des souches pouvant à moment donné présenter des caractères diploïdes.
- Organismes diploïdes totaux
La diploïdie concerne tous les gènes. (levures, champignons). Il existe des organismes qui possèdent 2 générations successives avec une alternance régulière haploïde / diploïde. Une génération haploïde, puis une génération diploïde résultant de la fusion de deux cellules différentes.
- Diploïdes partiels
Bactéries. La diploïdie est accidentelle (sous l’action de virus) ou résulte de phénomènes de parasexualité. Certains gènes seront introduits dans leur cellule, et ils vont se retrouver avec 2 gènes = diploïdes partiels.
Exemple théorique :
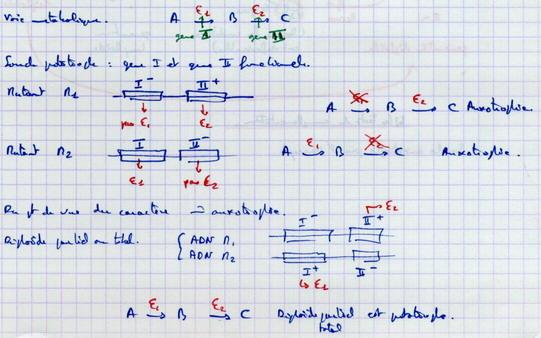
Dans ce cas, il y a complémentation. Les 2 génotypes se complètent. Il n’y a pas allélisme. On n’a pas deux fois le même allèle du même gène.

Il n’y a pas complémentation. Il y a allélisme.
I.C.2. Exemple de test d'allélisme complémentation : la levure
I.C.2.a. Leur cycle haplodiplophasique
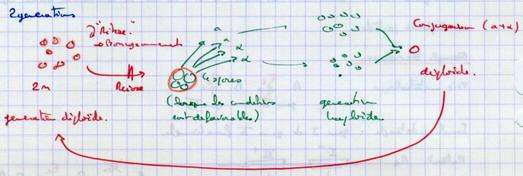
I.C.2.b. Test de complémentation
Souche auxotrophe pour l’uracile.
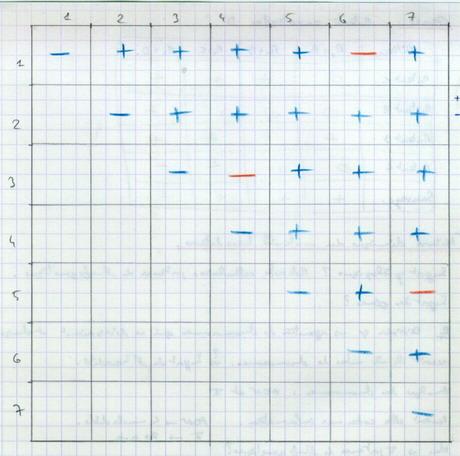
Souches haploïdes. + = complémentation ; - = pas de complémentation.
1-6 : pas de complémentation : mêmes gènes mutés : même souche.
3-4 : pas de complémentation : mêmes gènes mutés : même souche.
5-7 : pas de complémentation : mêmes gènes mutés : même souche.
4 gènes différents : (1-6), 2, (3-4), (5-7).
Conclusion : vraisemblablement, il faudra 4 enzymes (4 souches différentes).
![]()
Etude biochimique. Recherche du produit qui s’accumule anormalement par rapport à la cellule prototrophe (juste avant l’enzyme qui manque).Le test de complémentation permet de déterminer si les mutants sont identiques, ou sinon, le nombre de voies du métabolisme.
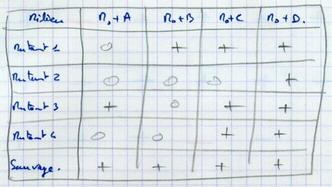
Exercice : Mutants auxotrophes D-.
II. Nature chimique du matériel héréditaire
Support cytologique ? Molécule cellulaire porteuse de l’information génétique. Quel est le support des gènes ?
Remarque : lors de la division cellulaire, il y avait apparition de chromosomes qui se divisaient et chaque cellule reçoit le même nombre de chromosomes. Les chromosomes sont-ils le support de l’hérédité ? L’analyse des chromosomes révèle qu’ils sont constitués d’ADN et de protéines. Ces molécules peuvent-elles coder une information ?
ADN → 4 nucléotides
π → 20 acides aminés
A vu du nombre de constituants, les protéines sont-elles porteuses de l’information génétique ?
II.A. Les étapes historiques : les travaux sur la transformation bactérienne
Ils ont commencé très tôt sur le pneumocoque.
II.A.1. Le matériel utilisé : les pneumocoques
Ils déclenchent la pneumonie. Cette bactérie est un diplocoque (formé de 2 cellules accolées). Il existe 2 catégories : virulente et non virulente.
Les souches virulentes sont lisses S avec capsule.
Les souches non virulentes sont détruites par les défenses immunitaires. Elles sont sans capsule = rugueuses.
Il existe des mutations qui font passer les souches d’une catégorie à l’autre.

II.A.2. La transformation bactérienne (Griffith, 1928)
Injection de bactéries de type S → mort des souris. La chaleur tue les bactéries virulentes. Il y a eu transformation bactérienne. Les bactéries R se sont transformées en bactéries S. |
 |
Occupation des capsides vides ? Non. Donc il existe un principe transformant qui serait thermostable venant des bactéries S et qui aurait pénétré les bactéries R pour leur conférer des caractéristiques de S. Il s’agit d’information génétique puisque les caractères modifiés étaient héréditaires. Il y avait eu échange d’information génétique. |
II.A.3. Identification du principe transformant Avery-MacLeod-MacCarty 1944
Reprise des expériences, mais in vitro.
II.A.3.a. Preuves indirectes que l'ADN est le principe transformant
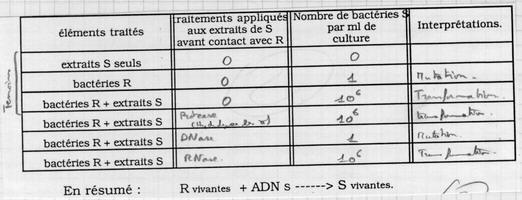
Le principe transformant de Griffith = ADN (preuve par la négative).
II.A.3.b. Preuve plus directe Avery
Obtention de clones : le principe transformant a modifié les caractères et ces modifications sont héréditaires. C’est donc l’information génétique = ADN. Avery a été surpris par ses résultats. Ce résultat a provoqué une certaine indifférence (comment coder avec seulement 4 nucléotides). Ce résultat était marginal pour lui. |
 |
II.B. Confirmation de ces résultats par des travaux sur les virus 1952
Travaux sur les virus car ils sont constitués d’ADN (ou d’ARN) et d’une capside protéique. Il s’agit donc d’un matériel de choix pour étudier l’ADN. Utilisation de la radioactivité.
II.B.1. Travaux sur le bactériophage Hersley et Chase 1952
Deux étapes :
- Préparation des virus
Dans le bactériophage, les protéines contiennent du souffre S absent de l’ADN. L’ADN, lui, contient du phosphore P, absent des protéines.
S* → Marquage des protéines → Capside*
P* → Marquage ADN*
On obtient donc 2 types de virus différents (radioactivité sur l’ADN ou sur la capside).
- Expérimentation
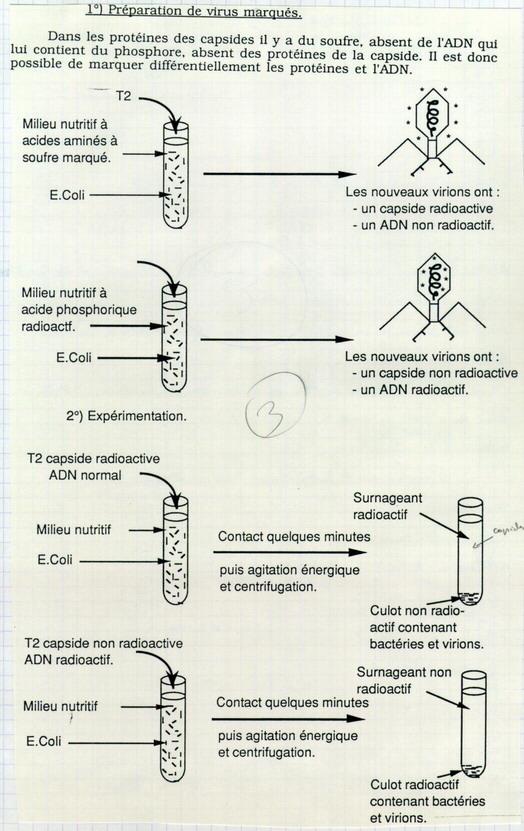
Interprétation : les nouveaux virions apparaissent dans le culot : dans les bactéries. Or les capsides ne pénètrent pas dans les bactéries. Seul l’ADN a pénétré à l’intérieur des bactéries. Donc, seul l’ADN peut être à l’origine des nouveaux virions. L’ADN seul permet de fabriquer de nouvelles molécules d’ADN et de nouvelles capsides. L’ADN porte l’information génétique du virus.
Conclusion : l’ADN est le support de l’information génétique. On s’intéresse alors à la molécule d’ADN. En 1953, Watson et Crick proposent le modèle moléculaire de l’ADN.
II.B.2. Travaux sur les virus à ARN
Virus de la Mosaïque du Tabac VMT.

Ce sont les acides nucléiques qui sont porteurs de l’information génétique (ADN ou ARN). Comment les acides nucléiques codent-ils l’information ? Comment cette information se réplique-t-elle identique à elle-même ? Comment apparaissent les mutations.
III. Réplication de l'ADN support de l'information génétique
L’information génétique contrôle les caractères des cellules. Ces caractères sont transmissibles de génération en génération. Donc l’information génétique se réplique identique à elle-même.
III.A. Le principe de cette réplication
III.A.1. Les différentes hypothèses envisageables
III.A.1.a. Réplication conservatrice
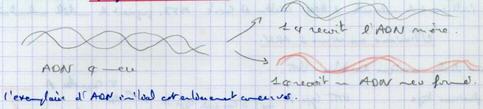
Il y fabrication d’un néo ADN.
III.A.1.b. La réplication semi conservatrice
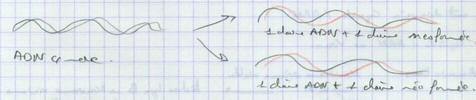
III.A.1.c. La réplication dispersive
Réplication plus aléatoire qui disperserait l’ADN initial de façon non régulière. Watson et Crick prédisent qu’il s’agit d’une réplication semi conservatrice en raison de la complémentarité des bases : modèle prédictif. |
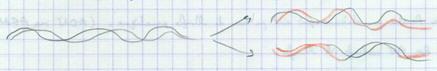 |
III.A.2. Les preuves expérimentales de la réplication semi conservatrice Meselson Stalh 1956
Technique de centrifugation en gradient de concentration.
III.A.2.a. Principe de l'expérimentation
Solution de saccharose ou de chlorure de césium. Centrifugation → Sédimentation selon un gradient de concentration régulier. On ajoute dans le tube l’ADN à centrifuger.
On obtient la séparation des ADN de masses différentes. La technique de détection utilise les propriétés d’absorption des UV par les cycles.
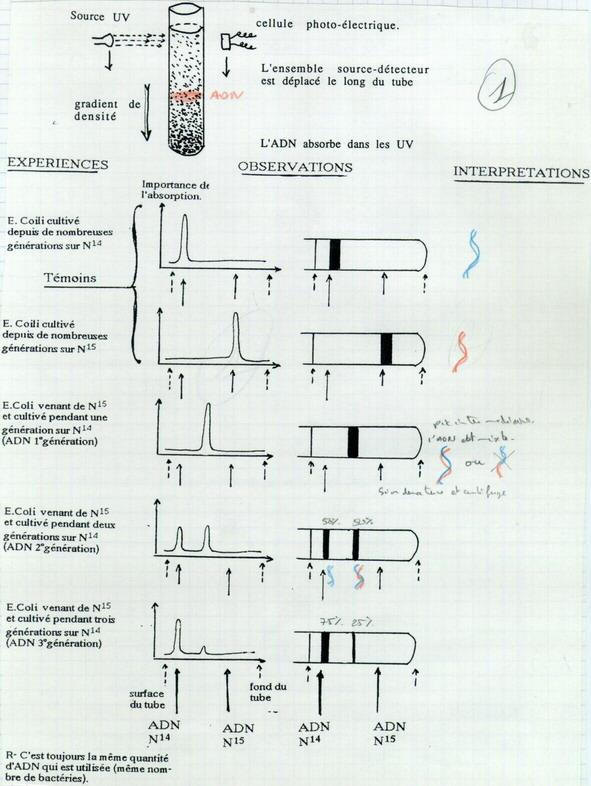
III.A.2.b. Résultats expérimentaux
Culture bactérienne, séparation de l’ADN et centrifugation.
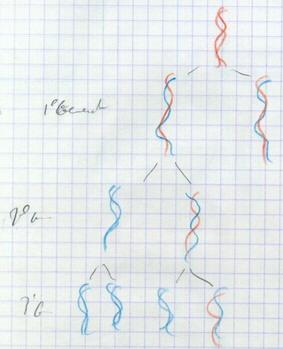
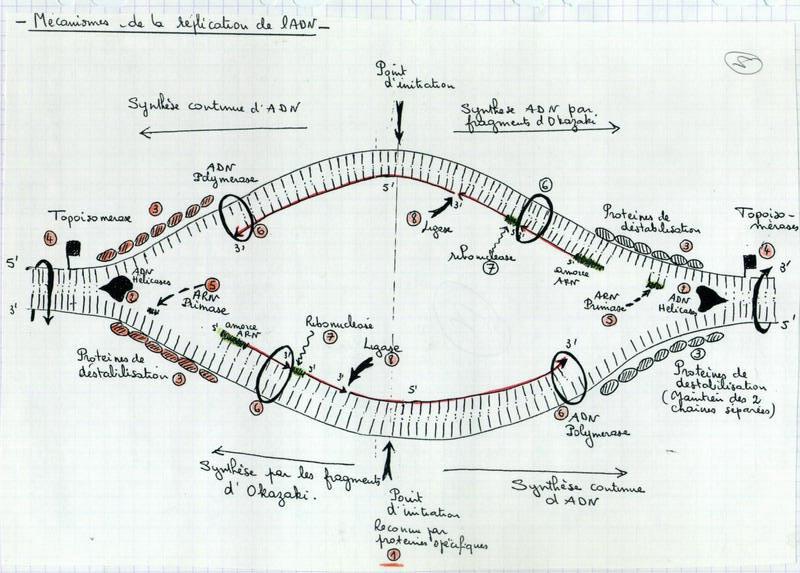
III.B. Mécanismes de cette réplication
III.B.1. Cette réplication est basée sur la complémentarité des bases azotées
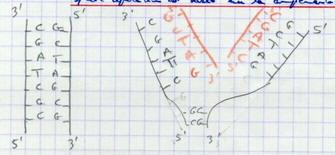
III.B.2. Point d'initiation de la réplication
- Unique chez les procaryotes et les virus (Observations Cairn)
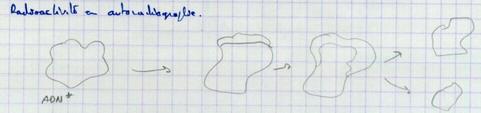
Donc le point d’initialisation est unique.
- Multiple chez les eucaryotes
| S’explique car les chaînes sont plus longues. La réplication est plus rapide si elle est commencée en plusieurs endroits. |  |
III.B.3. La synthèse est-elle unidirectionnelle ou bidirectionnelle ?

III.B.4. Principe général de l'élongation

III.B.5. Conditions d'action de l'ADN polymérase
- Sur extrémité 3’
- Sur matrice
- Si déjà commencé – nécessité d’une amorce
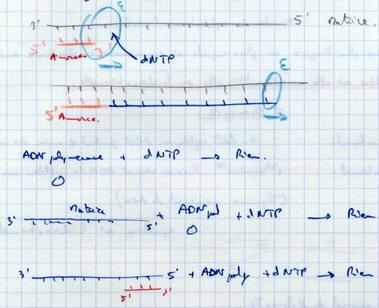
Conditions nécessaires : Matrice + Amorce + Extrémité 3’ libre.
III.B.6. Comment se fait le synthèse sur le brin 3' → 5'
Problème : La synthèse se fait sur les 2 brins.
|
 |
|
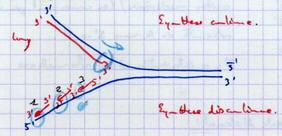 |
III.B.7. Qu'est-ce qui sert d'amorce ?
III.B.7.a. Observations
Lorsque les fragments d’Okazaki sont centrifugés par gradient de concentration, ils ont une densité intermédiaire entre les ADN et les ARN (mixtes).
La réplication nécessiterait des dNTP et des NTP, des ADN et des ARN.
Si on met de l’ARNase, on n’a pas de réplication de l’ADN. Il y a donc une étape qui nécessite de l’ARN.
Les enzymes qui bloquent les actions des ARN polymérases bloquent aussi la réplication de l’ADN.
III.B.7.b. Interprétation
On pense que l’amorce serait une amorce d’ARN fabriquée par une enzyme : Primase qui prend l’ADN pour matrice et peut fabriquer de petites chaînes complémentaires d’ARN sans avoir besoin d’amorce. Elle sera libérée au bout d’un certain temps.
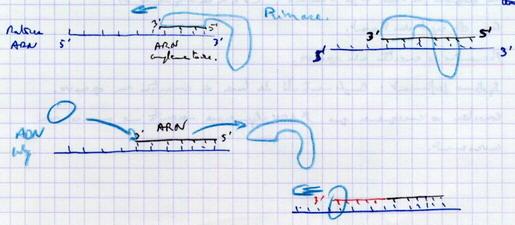
III.B.8. L'élimination des amorces d'ARN
L’ADN polymérase possède aussi une action exonucléasique (capable d’hydrolyser des acides nucléiques en commençant par une extrémité dans le sens 5’ → 3’. Il y a intervention d’une ribonucléase (hydrolyse les ARN). |
 |
III.B.9. La liaison des fragments d'Okazaki
Lorsque les amorces sont grignotées, on a 2 fragments d’Okazaki au contact l’un de l’autre sans être liés. |
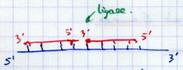 |
III.B.10. Il y a correction des erreurs de réplication
Fondamental pour la conservation de l’information génétique. Elle s’arrête si erreur (besoin d’une amorce). L’ADN polymérase va hydrolyser le dernier nucléotide qui n’est pas à se place pour l’éliminer (retour en arrière). Réaction exonucléasique 3’ → 5’. L’enzyme peut grignoter par l’avant et par l’arrière (3 sites catalytiques différents). Puis elle repart vers l’avant. |
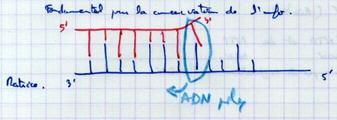 |
III.B.11. Problème d'ouverture de la double hélice lors de la réplication
III.B.11.a. La séparation des deux chaînes
Elle se fait grâce à une protéine : l’Hélicase. C’est une protéine allostérique qui utilise de l’ATP pour changer de conformation. Il y a des tensions physiques pour séparer les 2 chaînes. |
 |
III.B.11.b. Maintien des 2 chaînes séparées
Les chaînes ont tendance spontanément à se ré-apparier pour reformer la double hélice. Dès que l’hélicase agit, des protéines viennent se fixer (protéines de déstabilisation) et maintiennent les chaînes séparées.
III.B.11.c. Problème du désenroulement de l'ADN

Quand la tension est relâchée, les deux brins séparés sont liés de nouveau et l’enzyme de relaxation est libérée.
Problème énergétique ? Non car fonctionne quasiment à énergie constante.
Il existe 2 types d’enzymes qui se fixent respectivement sur 1 ou 2 brins.
Modèles.
III.B.12. Les ADN polymérases
Très grosses molécules qui possèdent plusieurs sites actifs.
- Fixation sur la matrice
- Fixation sur l’amorce (antiparallèle)
- Reconnaissance de l’extrémité 3’
- Fixation désoxyribonucléotide triPhosphate (en présence de Mg)
- Catalytique exonucléasique 5’ → 3’ à l’avant
- Catalytique exonucléasique 3’ → 5’ à l’arrière (correction sur épreuve)
- Site de vérification du bon appariement (à l’arrière)
Chez les procaryotes, il y a 1 seul type d’ADN polymérase. Chez les eucaryotes, il y a 2 types d’ADN polymérases (1 sur le brin précoce pour la synthèse en continu et une autre sur les brins tardifs [fragments d’Okazaki]).
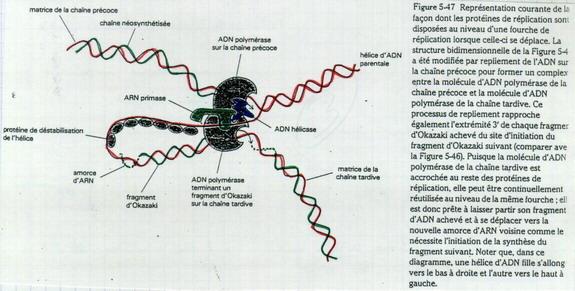
| Système compliqué et coûteux en ATP. Pourquoi un enchaînement 5’ → 3’ ? Si sens 3’ → 5’ la correction sur épreuve est impossible. |
 |
Sujets
- Les protéines de la réplication
- Réplication : mécanisme haute fidélité
- Intérêt d’une information génétique en double chaîne
IV. Modifications de l'information génétique : les mutations
Modifications héréditaires des caractères. Modification transmissible = affecte le matériel génétique (ADN).
IV.A. Les différents types de mutations
IV.A.1. Mutations ponctuelles (qui n'affectent qu'une paire de bases)
IV.A.1.a. Substitution d'une paire par une autre
- Transitions : A-T remplacé par C-G
- Transvertion : A-T remplacé par T-A
IV.A.1.b. Délétion
Disparition d’une paire de bases. Tout le code génétique est décalé. Les conséquences sont bien plus importantes.
IV.A.1.c. Addition d'une paire de bases
Décale tout le code génétique d’une base.
IV.A.2. Les mutations géniques 10-100 bases
Affectent un nombre important de paires de bases. Toute une partie du gène est modifiée. Il s’agit souvent de délétions qui provoquent un blocage du fonctionnement du gène.
IV.A.3. Mutations chromosomiques
Affectent l’ensemble des chromosomes
IV.A.3.a. Délétions
Chromosomes plus courts. Visible lors de caryotypes. |
 |
IV.A.3.b. Translocations
Certains gènes sont déplacés. Perturbation du phénotype. |
 |
IV.A.3.c. Inversions
Conséquences au plan métabolique. |
IV.A.4. Mutations génomiques
Tout le génome est modifié. Aspect quantitatif → qualitatif (caractères modifiés).
IV.A.4.a. La monoploïdie
L’individu est haploïde au lieu d’être diploïde.
IV.A.4.b. La polyploïdie
Tout est multiplié : 3n, 4n, … Triploïdie, tétraploïdie…
Ce phénomène est fréquent chez les plantes.
IV.A.4.c. Aneuploïdie
La formule génétique est anormale.
- Monosomie
2 x 22 + X monosomie au niveau des gonosomes.
- Trisomie
Trisomie 21 : 3 x 21 syndrome du mongolisme 47 chromosomes (le chromosome 21 est présent en 3 exemplaires).
IV.B. Modifications ponctuelles des séquences d'ADN : mutations ponctuelles
IV.B.1. Modifications spontanées
IV.B.1.a. Dépurinations
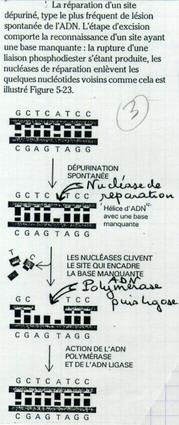
Ablations de bases puriques A et G dans la molécule d’ADN. Il manque 1 base. C’est accident survient avec une fréquence de 5 000 fois par jour et par cellule. S’il n’est pas réparé, il pose problème lors de la réplication et lors de l’expression du gène. |
IV.B.1.b. Désamination
Certaines bases azotées peuvent être désaminées (modifiées).
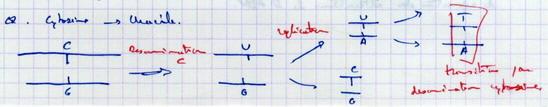
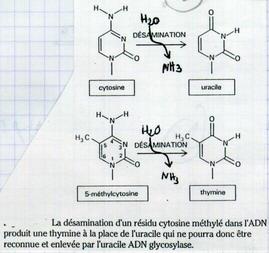
100 bases par et jour et par cellule sont désaminées.
IV.B.2. Modifications provoquées par des agents externes
IV.B.2.a. Formation de dimères de Thymine par les UV
Ces deux bases se lient entre-elles. Problèmes au niveau de la réplication et de l’expression. Compte pour 1 → décalage du code. |
 |
IV.B.2.b. Transitions induites par l'acide nitreux HNO2
Va modifier les bases : exemple Adénine → Hypoxanthine Le matériel génétique est modifié très souvent. Malgré cela, les mutations sont rares car il existe des phénomènes de réparation des altérations de l’ADN. |
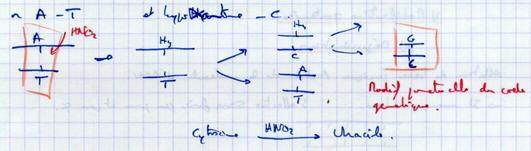 |
IV.C. Les phénomènes de "réparation" de l'ADN
IV.C.1. Principe général
La réparation ne peut intervenir qu’à un stade précoce : une seule base modifiée (quand la base en face n’a pas été modifiée = matrice correcte). Enzymes de reconnaissance et d’excision (hydrolysent les liaisons phosphoester).
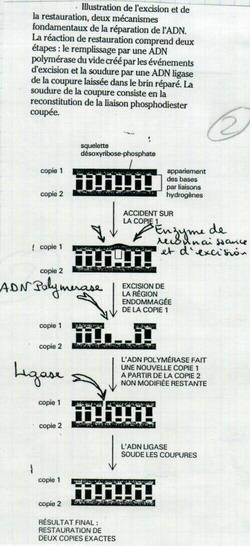
IV.C.2. Dans le cas d'une dépurination
Les enzymes repèrent l’absence d’une base. Elles excisent de part et d’autre. Puis l’ADN polymérase, puis ligase.
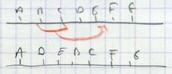
IV.C.3. Réparation d'une modification de Cytosine en Uracile

Conclusion
Les altérations peuvent être fréquentes. La cellule possède un équipement enzymatique et protéique destiné à la réparation de ces lésions. Ces réparations sont possibles car il existe 2 chaînes. Elles demeurent possibles tant que la matrice complémentaire est conservée. Si les 2 chaînes sont modifiées, impossibilité d’intervenir pour réparer. Cette « maintenance » de l’ADN assure le caractère de l’espèce.
Il existe des mutations par déficience : il manque les enzymes pour éliminer les dimères de Thymine = déficience génétique → tumeurs de la peau.
Sujets
- Pourquoi y a-t-il si peu de mutations (différents types et réparation) ?
- Mécanismes assurant la conservation de l’information génétique de génération en génération. Fidélité réplication (semi conservatrice), correction sur épreuve + réparation des altérations.