
Introduction
Les différences observées se situent au niveau de l'organisation et de la localisation du matériel génétique.
I. Localisation et structure du matériel génétique chez les eucaryotes
I.A. Le matériel génétique est localisé dans le noyau
Le noyau est délimité par une double membrane qui le différencie du Réticulum Endoplasmique. Cette membrane est percée de pores permettant la communication noyau / hyaloplasme (différence par rapport aux mitochondries et aux chloroplastes).
Intérieurement, cette membrane est doublée par la lamina nucléaire : c'est une couche protéique qui double la membrane intérieurement. Sur cette lamina est fixée la chromatine. Sur la membrane externe, il peut y avoir des ribosomes fixés.

On constate la présence de nombreux pores nucléaires bordés de protéines et d'une protéine centrale. A l'intérieur du noyau se trouve la chromatine, un ou plusieurs nucléoles (synthèse des ARN ribosomaux, maturation) (association protéines-ARNr = Sous Unités ribosomales).
Le reste constitue le nucléoplasme : eau, Sels Minéraux, protéines, ARNt, ARNm, pré messager qui communique avec le hyaloplasme par les pores nucléaires. Cette structure disparaît au moment de la division cellulaire : la membrane disparaît, le nucléole aussi, et la chromatine se condense en chromosomes
I.B. Structuration du matériel génétique (par rapport aux procaryotes)
I.B.1. Aspect
Il se présente sous la forme de molécules linéaires d'ADN et se trouve très souvent en deux exemplaires identiques (mêmes gènes, mais pas forcément les mêmes allèles) pour les organismes diploïdes. Il existe aussi des eucaryotes haploïdes.
I.B.2. Les gènes peuvent être répétés
On peut trouver plusieurs exemplaires d'un même gène.
I.B.2.a. Mise en évidence
Étude des phénomènes de renaturation : on extrait l'ADN des cellules, on le fragmente (enzymes de restriction), on le dénature par chauffage et on obtient deux chaînes d'ADN monocaténaire.
Puis on refroidit : il y a renaturation, chaque chaîne rencontre la chaîne complémentaire. La vitesse de ré association augmente avec la probabilité de rencontre. On distingue 3 grandes catégories d'ADN :
- ceux qui se renaturent très vite
ces fragments existent en de très nombreux exemplaires : ce sont des gènes fortement répétés. - ceux qui se renaturent moins rapidement
ce sont des gènes moyennement répétés - ceux qui ne se renaturent qu'à la fin
ce sont des gènes uniques ou non répétés.

I.B.2.b. Les différentes classes de gènes
- Les gènes fortement répétés : séquences
Ce sont des gènes présents à plusieurs milliers ou millions d'exemplaires. Ils représentent 10 à 25 % du génome. Ces sont des séquences très courtes, groupées en bloc, essentiellement au niveau de l'hétérochromatine (très fortement condensée). Ce sont des zones où il n'y a jamais de transcription. Leur rôle est mal connu. Les cellules possèdent 10 à 25 % d'ADN qui apparemment ne sert à rien.
- Les gènes moyennement répétés (plusieurs centaines)
- Les gènes qui codent pour les ARN ribosomiques et de transfert. Ce sont des gènes transcrits mais pas traduits. Le phénomène d'amplification ne s'effectue qu'à un niveau : la répétition des gènes constitue le seul niveau d'amplification.
- Les gènes qui codent pour les histones de la chromatine
Le besoin est fort au moment de la réplication de l'ADN (division cellulaire). En très peu de temps, le nombre d'histones doit doubler. La présence de plusieurs exemplaires au niveau des gènes permet d'assurer rapidement le besoins de la cellule (chez l'homme : quelques dizaines d'exemplaires; chez l'oursin, environ 1000 exemplaires). - D'autres gènes dont le rôle est encore inconnu.
- Les gènes uniques (50 % du génome)
Ils sont présents en 2 exemplaires (1 pour chaque chromatine homologue). Ces gènes codent pour des protéines spécifiques, souvent métaboliques. Deux niveaux d'amplification, la transcription et la traduction.
I.B.3. Les gènes sont morcelés : gènes mosaïques 1977
Les ARN messagers sont plus courts que les fragments d'ADN qui servent à leur formation. Exemple du gène de l'ovalbumine dans une cellule oviducte d'oiseau.
Isolement de l'ARNm (ovalbumine). L'ovalbumine représente 75 % de la synthèse des protéines. Fragmentation du génome des cellules de l'oviducte. Dénaturation de l'ADN et mise en présence de l'ADN monocaténaire et de l'ARNm avec repérage par radioactivité. On observe des figures surprenantes :

L'ARNm est plus court que l'ADN qui a servi de matrice à sa synthèse. L'ARNm est plus court que le gène or la transcription est un phénomène continu. Et l'ARNm sert à la synthèse des protéines. Il comporte donc l'intégralité de l'information génétique nécessaire à la synthèse. Le gène possède donc l'intégralité de l'information génétique mais ses séquences ne sont pas continues : l'information génétique est discontinue.
On a appelé introns les séquences d'ADN non porteuses d'information génétique intercalés dans la partie codante (exon).
L'information génétique sur l'ADN eucaryote est morcelée. Les gènes sont en mosaïque : alternance de parties non codantes et de parties codantes.
Chez les procaryotes on distingue :
- Archébactéries : bactéries ancestrales (les plus anciennes)
Chez les archébactéries, les gènes sont en mosaïque - Eubactéries : les plus nombreuses et les plus évoluées
Chez les eubactéries, les gènes ne sont pas morcelés : l'information génétique est continue. Les eubactéries sont très évoluées, elles ont perdu les introns au cours de l'évolution (gain de place dans la cellule, économie lors de la réplication, donc augmentation des performances)
Les eucaryotes trouvent donc leurs racines chez les archébactéries. On a trouvé des introns dans les mitochondries, ce qui les rapproche des archébactéries.
II. Traits distinctifs au niveau de la transcription
II.A. Structure des gènes
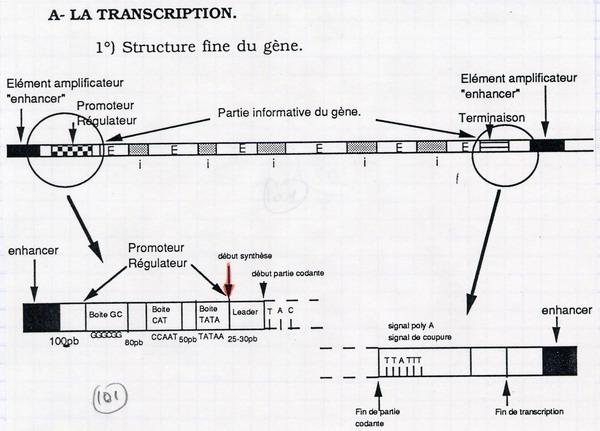
II.B. Les ARN polymérases
Chez les procaryotes, il existe une seule ARN polymérase. Chez les eucaryotes, il existe 3 ARN polymérase différentes. Mise en évidence car les poisons n'ont pas le même effet sur les 3.
- La polymérase 1 catalyse la bio synthèse des ARN pré ribosomiques dans le nucléole
- La polymérase 2 synthétise les ARN pré messagers
- La polymérase 3 synthétise les ARNt les ARN ribosomaux légers 5S à l'extérieur du nucléole et les petits ARN du noyau (Sn ARN = small nuclear ARN)
Ces trois enzymes différentes sont régulées par trois mécanismes indépendants.
II.C. Les étapes de la transcription
II.C.1. L'initiation
Dans le promoteur, on retrouve des séquences très conservées (presque communes à tous les promoteurs) : Boite CG, Boite CAT, Boite TATA. L'ARN polymérase se positionne sur le promoteur :

Intervention d'une protéine régulatrice : le facteur TATA. L'ARN polymérase ne se fixe sur le promoteur que si le facteur TATA est en place, et il faut qu'un facteur d'initiation intervienne pour la dissociation des deux chaînes.
II.C.2. L'élongation
II.C.3. La terminaison
Signal de terminaison : beaucoup de thymine → adénies = queue poly A.

Le résultat de la transcription est un pré ARN messager. Il n'y a pas encore eu l'excision des introns.

II.C.4. Images de la transcription


Chromosomes après la duplication.
Les chromatides ne sont pas séparées. Chromosome en écouvillon (pour nettoyer les bouteilles)
Zone où il n'y a pas de transcription ; chromomère inactif = hétérochromatine.
Chromomère actif = euchromatine.
Sur l'illustration, "poils" d'ARNm en synthèse : les molécules d'ARN polymérase fonctionnent les unes derrière les autres. C'est un phénomène d'amplification.
III. Maturation des ARN
III.A. Maturation des ARNr
Les ARNr 45S sont découpés en petits morceaux (cf. Procaryotes).
III.B. ARN de transfert
| Maturation analogue à celle des ARNt des procaryotes. Excision de fragments d'ARN en bout de chaîne ou intrachaine. Ex : Pré ARNt Thyrosine : |  |
III.C. Maturation des ARN messagers
Chez les eucaryotes, ARN monocistronique. La maturation s'effectue dans le noyau.
III.C.1. Allongement de la queue poly A en 3'
Grâce à une enzyme, la polyadénylate polymérase : allongement sur l'extrémité 3' d'adénine à partir d'ATP. Pas besoin de matrice pour agir, seulement d'une extrémité 3'. Cette queue poly A est nécessaire à la stabilité des ARNm. Dans le hyaloplasme, elle est dégradée et l'ARNm se désorganise. Cette queue poly A est nécessaire au passage à travers les pores nucléaires. Elle est nécessaire pour isoler les ARNm sur des colonnes constituées de poly T (résine).
III.C.2. Modification de l'extrémité 5' : formation du 5' Cap

Cette modification est nécessaire pour que l'ARNm soit traduit sur le ribosome. Si elle n'est pas présente, la traduction n'est pas initiée : la petite Sous Unité ne se fixe pas sur l'ARNm. La fixation du 5' Cap se fait très tôt : dès le début de la transcription, grâce au facteur d'élongation. Ce facteur d'élongation est nécessaire à la transcription de tout ARNm. S'il est absent, tout l'ARNm est dégradé au cours de la transcription. Le 5' Cap est nécessaire à la préservation de la dégradation des ARNm.
III.C.3. L'excision des introns
| Au moment de la transcription, il y a des introns et des extrons. Les introns, qui sont inutiles, seront excisés. Intervention de protéines du noyau associées à de petits ARN : les snARN → snRNP. |  |
| L'excision va se faire depuis le site donneur jusqu'au site accepteur. Les extrémités des introns sont constituées de bases quasi identiques à peu près pour tous les introns. Les snRNP vont se fixer sur les extrémités de l'intron. On observe un repliement de l'intron. |  |
Association quaternaire de snRNP qui forment une liaison covalente avec une zone un peu plus éloignée du site accepteur. Coupure au niveau du site donneur, coupure au niveau du site accepteur et réaction de liaison à l'intérieur de l'intron, puis libération de l'intron sous forme d'une boucle : lasso associé aux snRNP.
 |
 |
C'est un phénomène extrêmement précis : décalage d'une seule base et tout le codage est faux. Pourquoi les petits ARN sont-ils associés aux protéines ? Ces petits ARN se lieraient avec les extrémités des introns pour former une boucle. Séquences complémentaires des extrémités des introns ? Ce phénomène existe également dans le cas des ARN ribosomaux sauf qu'il y a coupure précise mais pas épissage (re-soudure = liaison covalente).
Pour les ARNr, chez certains protozoaires, et pour les ARN messagers des mitochondries de levures, les phénomènes de coupure et d'épissage (élimination des introns) se faisaient sans le concours des snRNP, c'est à dire sans enzymes. Les introns possédaient un pouvoir catalytique sur leur propre excision. Ils agissent comme une enzyme. On parle de ribozyme = fragment d'ARN à pouvoir catalytique.
Evolutivement, l'excision des introns se faisait sans le concours des protéines. Cette action catalytique des ARN est à rapprocher de l'extrême richesse des ARN dans les ribosomes. Si les ribosomes sont riches en ARN, alors c'est que les ARN ont un rôle prépondérant, et peut être un pouvoir catalytique dans la traduction.
Arguments : l'analyse des ARN ribosomaux montre des séquences très constantes (conservées) d'un organisme à l'autre alors que les protéines du ribosome sont très différentes : les protéines n'y jouent donc pas un rôle fondamental. L'important, c'est l'ARN et son pouvoir catalytique. La puissance catalytique des protéines vient de leur conformation 3D. Or les ARN ont aussi une conformation 3D qui fait apparaître des sites actifs : couplage de bases complémentaires en disposition inverse qui est à l'origine de formes complexes qui constituent les sites catalytiques.
III.D. Cas particuliers de maturation : modification post transcriptionnelle
III.D.1. Epissages différentiels
Certains pré ARNm pourraient subir des excisions d'introns différentes. Exemple des protéines fobronectines qui jouent un rôle dans l'adhérence cellulaire. Il existe moins de gènes que de fibronectines différentes. Un gène donne un ARN pré messager :

Les phénomènes d'excision par épissage différentiel sont régulés. A partir d'un même ARN pré messager, on obtient 3 protéines différentes.
1 gène → 3 protéines : la notion 1 gène → 1 protéine est discutée.
III.D.2. Phénomènes d'édition
Il existe des protéines pour lesquelles on ne trouvait pas le gène. Exemple chez le Trypanosome (protozoaire) parasite de l'homme et responsable de la maladie du sommeil. La Sous Unité III de la cytosome oxydase est une protéine. On connaissait la séquence de la protéine, et on s'est mis à rechercher le gène. Mais le gène est extrêmement différent de l'ARNm : 357 nucléotides de plus dans l'ARNm dispersés sur 121 sites différents !
Après transcription, on obtient un pré ARN. Il y a ensuite des phénomènes d'insertion de nucléotides supplémentaires : le code porté par le gène est modifié ! L'information génétique est transcrite, puis modifiée. Ce phénomène est répandu à d'autre cellules. Dans ces cellules :
- Insertion de nucléotides
- Elimination de nucléotides (ponctuelles, 1par 1)
- Substitutions de nucléotides d'un codon jusqu'à un codon stop.
Ce sont des phénomènes régules qui aboutissent à une protéine fonctionnelle. Un gène soumis à plusieurs phénomènes d'édition donne plusieurs protéines.
Chez les eucaryotes, pour la traduction, il y a une dissociation dans l'espace : la transcription se fait dans le noyau, alors que la traduction s'opère dans le hyaloplasme. Chez les procaryotes, ces phénomènes sont simultanés.
IV. Maturation des Protéines
Les protéines fabriquées sur les ribosomes ne sont pas fonctionnelles. Une protéine ne devient fonctionnelle que lorsqu'elle a acquis sa configuration tridimensionnelle. Cette configuration ne sera atteinte qu'après certaines modifications.
IV.A. Excisions d'acides aminés
IV.A.1. Excision de l'extrémité N terminale
La traduction commence par la mise en place de la Méthyonine (Formylméthyonine) codon AUG. Or, cette méthyonine n'existe plus dans la protéine définitive : il y a un phénomène d'excision : le premier acide aminé est hydrolysé.
IV.A.2. Excision de la séquence de signalisation peptid signal
Lorsqu'une protéine est fabriquée sur les ribosomes, elle peut avoir plusieurs destinations possibles :
- Hyaloplasme (ex : glycolyse )
- Réticulum Endoplasmique Rugueux → Vésicules (lysosomes ou exportations : enzymes, hormones, neurotransmetteurs)
- Membrane (Protéines membranaires)
La destination est différente selon leur nature. On sait pourquoi certaines entrent dans le RER. Ce qui permet l'orientation de la la protéine, c'est la 1ère séquence d'acides aminés = Peptid Signal. Il s'implante dans une ribophorine qui s'implante sur le réticulum des ribosomes. Les ribosomes sont situés au niveau de protéines canaux. La protéine va passer dans le RE. Puis une enzyme hydrolyse le signal qui n'est plus utile : peptidase du signel. La séquence de signalisation permet l'orientation de la protéine vers sa destination.
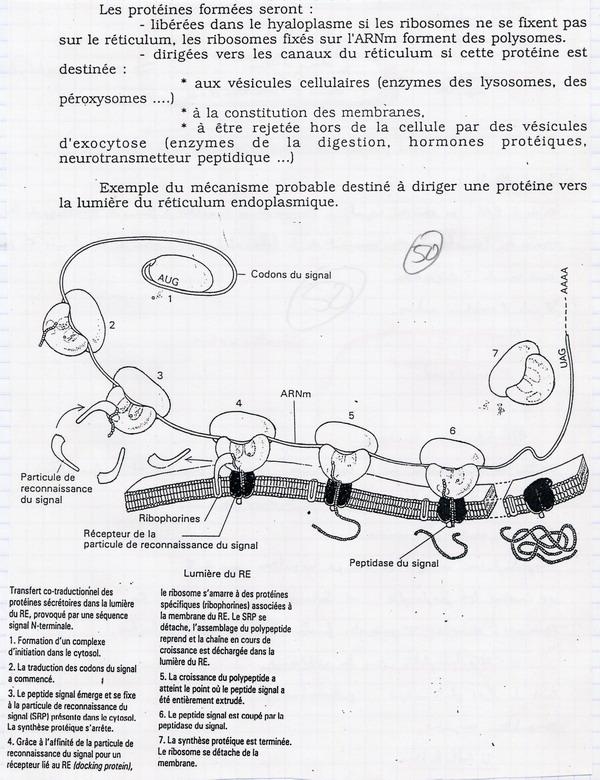
IV.A.3. Excisions intra moléculaires
Elles sont nécessaires pour rendre la molécule active. Elles ne sont pas suivies d'un épissage. La molécule initiale donne 2 ou 3 molécules qui restent fixées par des ponts Di-Sulfures. Exemple : enzymes, chimiotripsine, insuline.
Insuline : Hormone hypoglycémiante produite par les cellules β des îlots de langerans du pancréas :

| 3 ponts Di-Sulfures La protéine va dans le RER où le peptid signal est excisé. La pro insuline est inactive : c'est une forme de stockage dans les vésicules (où elle est inactive). Au moment de la libération, l'excision de la chaîne C est réalisée par excision de de dipeptides. Il reste l'insuline active. |
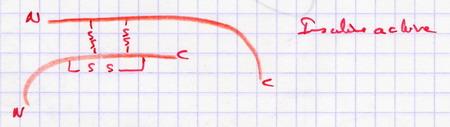 |
IV.B. Modifications chimiques
Fixation de groupements ou de radicaux qui entraîne une modification des propriétés et des conformations.
IV.B.1. Phosphorylation des radicaux hydroxydés
Exemples : Thyrosine, sérine, Thréonine...
La phosphorylation apporte des charges négatives à la protéine. Par exemple, la caséine est très phophorylée, ce qui provoque le phénomène de saturation par les ions calcium (Ca2+).
La phosphorylation est nécessaire à certaines protéines pour être fonctionnelles : glycogène phosphorylase...
IV.B.2. Carboxylation sur les radicaux
Exemples de la carboxylation de l'acide glutamique, de la prothrombine qui intervient dans la coagulation sanguine.
IV.B.3. Méthylations
IV.B.4. Glycosylations
Des chaînes polysaccharidiques sont fixées sur la protéine, qui devient une glycoprotéine. Elles se font dans le RER et dans l'appareil de Golgi.
IV.B.5. Addition de groupements prostétiques
Nécessaire pour rendre certaines protéines actives : hémoglobine, myoglobine, cytochromes : Noyau tetrapyrolique + Fer.
IV.B.6. Formation de ponts disulfures intrachaîne
Modification chimique de nature covalente qui stabilise une structure 3D.

Chez les eucaryotes, les phénomènes sont plus complexes, plus lents et plus longs.
V. Régulation de l'expression génétique chez les eucaryotes
V.A. Caractéristiques générales
V.A.1. Le modèle de Jacob et Monod ne s'applique pas
Chez les eucaryotes, les gènes d'une même voie métabolique sont dispersés sur plusieurs chromosomes : pas d'induction ou de répression coordonnées. Les gènes ne sont pas regroupés en opérons sous contrôle d'un même promoteur.
V.A.2. Régulation plus lente
Les ARNm ont une durée de vie plus longue, mais ce n'est pas un obstacle : les cellules eucaryotes vivent dans des milieux dont les caractéristiques varient peu. Elles n'ont pas besoin de s'adapter à des changements brusques du milieu.
V.A.3. Ce n'est pas la même partie du génome qui s'exprime dans les différents tissus
Gènes domestiques : ce sont des gènes qui s'expriment dans toutes les cellules de l'organisme (ex : enzyme de la glycolyse...). D'autres gènes, spécifiques, ne s'expriment que dans certains tissus. Par exemple, les gènes qui assurent la synthèse de l'urée ne s'expriment que dans les cellules hépatiques. Dans les cellules musculaires, les gènes responsables de la synthèse de l'actine et de la myosine sont très actifs.
La régulation de l'information génétique devient un problème de différenciation cellulaire car dans toutes les cellules, c'est la même information génétique qui est présente. Dans certaines cellules, certains gènes s'expriment et pas d'autres.
V.B. Problème de différenciation cellulaire
V.B.1. La différentiation cellulaire cytoplasmique est souvent irréversible
Si la cellule a atteint un degré de spécialisation important, cette cellule perd la possibilité de se diviser et est incapable ensuite d'évoluer dans une autre voie. La différenciation est irrémédiable.
Exception : les cellules végétales : elles peuvent se dédifférencier puis se redifférencier dans une autre voie.
Chez les animaux supérieurs, ce phénomène n'existe pas.
V.B.2. Au niveau nucléaire elle n'est pas irréversible
On prélève une cellule intestinale de têtard. On en prélève le noyau et on l'injecte dans un oeuf non fécondé dont on a détruit le noyau. Le noyau de la cellule intestinale possédait toutes les potentialités pour la création d'un être vivant complet. Le noyau possédait encore tous les gènes. Les gènes réprimés ne le sont pas de manière définitive. Il peuvent s'exprimer dans un contexte différent. La réaction n'est pas irréversible. On n'a pas de modèle unique pour la régulation chez les eucaryotes.
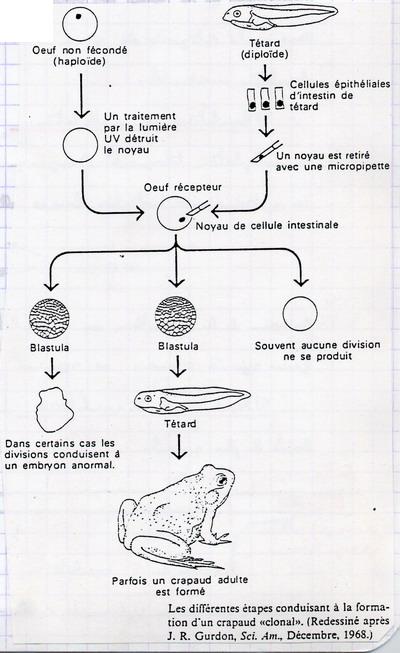
V.C. Quelques exemples de régulation
V.C.1. Amplification des gènes
V.C.1.a. Exemple : cellule ovocyte de vertébré.
Il doit accumuler très rapidement des protéines de réserve, donc il a besoin de ribosomes. Or cet ovocyte est capable en quelques semaines de fabriquer 1012 ribosomes alors qu'une cellule normale n'en fabrique que 105. La nécessité d'ARN ribosomaux se traduit par une amplification des gènes ribosomaux. Dans la cellule, on observe de petits ADN circulaires qui contiennent les gènes ribosomaux. Ces ADN vont se multiplier. Une chaîne est cassée, se déroule et l'ADN polymérase entre en action. On obtient 1000 fois plus de gènes ribosomaux. On ne sait pas comment apparaît le premier gène circulaire.

V.C.1.b. Exemple des cellules folliculaires de drosophile
Elles fabriquent des protéines formant la coque dans les oeufs, ce qui nécessite une duplication des gènes.


V.C.2. Régulation par perte d'information génétique
Ce sont des phénomènes assez exceptionnels : Ascaris ou copépodes.
Dans certaines cellules, on assiste à une élimination d'ADN : réduction de la taille des chromosomes : élimination d'une grande quantité d'information génétique. Une seule catégorie de cellules n'élimine pas d'ADN, il s'agit de la lignée reproductrice. Ce type de régulation est irréversible.
V.C.3. Régulation par méthylation
Les méthylations bloquent l'expression des gènes. Les cytosine sont méthylées lorsqu'elles sont dans cette disposition : Au cours de la réplication, on assiste à la méthylation de la nouvelle cytosine. Les gènes réprimés dans la cellule mère le seront dans la cellule fille. |
 |
V.C.4. Régulation par protéines régulatrices
Modèle le plus général : Il peut y avoir un repliement de la molécule d'ADN qui va placer l'enhancer au dessus du promoteur et les protéines régulatrices sur l'enhancer vont pouvoir réguler le promoteur. |
 |
| Provoque la libération d'histones au niveau de la boite TATA. Puis fixation du facteur TATA... Intervention de très nombreuses protéines régulatrices. | 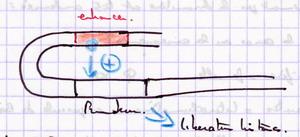 |
V.C.5. Régulation par des facteurs extra cellulaires
Par des hormones, comme par exemple les stéroïdes. Exemple de l'ecdysone qui diffuse à travers la membrane lors de la mue. Le complexe ecdysone - récepteur migre vers le noyau vers les chromosomes géants ou polyténiques. Ce sont des chromosomes dans lesquels il y a eu des duplications d'ADN qui n'ont pas été suivies de séparations : filaments d'ADN accolés les uns aux autres. Le complexe ecdysone - récepteur est à l'origine d'un puff sur le chromosome. La chromatine se déploie pour que la transcription se fasse. Il y a formation d'une protéine qui va inhiber le gène qui vient d'être activé (parricide) et qui stimule un autre gène (puff tardif). Il s'agit s'un système en cascade. La mécanique est amorcée. Une substance extra cellulaire agit sur l'expression des gènes. |
 |
V.C.6. Régulation et hétérochromatine
Lorsque les gènes ne s'expriment pas, la chromatine est très condensée : hétérochromatine. Cette condensation est-elle la cause ou la conséquence ? Cas des chromosomes sexuels chez les femelles des mammifères
Un chromosome apparaît normal, et l'autre est condensé sous forme d'hétérochromatine = corpuscule de Barr. Un chromosome X est inactif. Ce peut être l'un ou l'autre, ce n'est pas toujours le même qui est inactif. Conséquences génétiques :

Ce phénomène se produit de manière aléatoire. On a l'impression qu'un chromosome lui suffit. Or, au cours du développement embryonnaire, les deux chromosomes X sont nécessaires et l'inhibition d'un chromosome X ne se produira qu'après le développement embryonnaire. Sauf dans la lignée germinale, le chromosome bloqué est bloqué définitivement.
V.C.7. Régulation par des phénomènes de transposition
Gènes sauteurs : transposons. Ce sont de petits fragments d'ADN qui peuvent quitter le chromosome pour s'intégrer ailleurs et bloquer le gène dans lequel ils s'intègrent. Ils peuvent ensuite s'exciser pour le rendre inactif.
V.C.8. Régulation au niveau de la maturation des ARN messagers
Exemple des épissages différenciés et des phénomènes d'édition.
V.C.9. Régulation au niveau de la traduction
V.C.9.a. Cas des ovocytes d'échinodermes
 |
 |
Bloquer la transcription n'a aucun effet : les ARN messagers étaient déjà présents dans l'ovule avant la fécondation (ils étaient alors bloqués). La traduction était alors bloquée. Le phénomène n'est pas régulé à la première étape. Problème de rapidite : permet d'accélérer les phénomènes qui suivent la fécondation pour conduire à l'embryon.
V.C.9.b. La synthèse de ferretine
Protéine qui permet de stocker le fer dans l'organisme.

En présence de fer, celui-ci vient se fixer sur la protéine régulatrice qui se libère :

Conclusion
Contrairement aux procaryotes, où il existe un petit nombre de modèles de régulations, chez les eucaryotes, il y a une très grande diversité des phénomènes de régulation de l'information génétique.
Lors de la différenciation cellulaire, elles sont définitives. Mais il existe d'autres régulations ajustées qui varient au cours de la vie de la cellule. Il n'y a pas de modèle unique de régulation. Le plus significatif est la régulation par les enhancer pour la régulation au cours de la vie de la cellule.